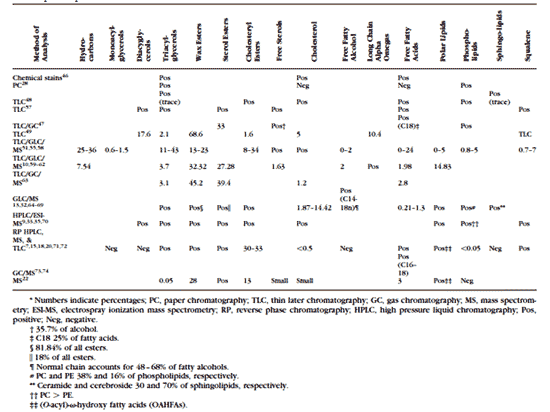
涙液層( TF: tear film )の分子構成(タンパク質と脂質など)と涙液層へのマイボーム腺の関与について解明することは、涙液層の不安定 化 、ドライアイ症候群、コンタクトレンズ( CL )不耐症および眼疾患の理解を深める上できわめて重要である。涙液層の脂質層は、水分の蒸発の抑制、 眼 表面の透明性の維持、細菌や塵埃・花粉など有機物から の眼の 保護といったさまざまな機能を持つ 1 。涙液層にはタンパク質、酵素、脂質、ムチン、塩類などの複雑な混合物が含まれており、それらによって涙液層の機能が発揮されている( Fig.1 )。研究者たちは、外層の脂質層は、 5 ~ 10 分子の厚みで、主としてワックスエステルとステロールエステルで構成されており、明確な連続する分子層形成するのではなく、互いに、そしてタンパク質とも 相互作用していると考えている 2,3 。干渉計を用いた研究によると、涙液層の油層の厚さは 20 から 160 nm である 4 。ここで脂質分子の大きさをおよそ 2.2 nm ( 22 Å )とすると、層ひとつの厚みは 11 から 44 nm という計算になる。極性および非極性の脂質が加わると脂質の厚みが増すことから、厚み の実測値と一致するためには、涙液層の脂質成分が多層の厚みを持っている、または別の寄与因子があることが考えられる 5 。
涙液層の不安定化の他覚所見と自覚症状は比較的よく把握されているのに対し、涙液層に特異的な成分分子、それらと本疾患および涙液層の不安定化との関係についての研究はまだ緒についたばかりである。本論説は、健常時と疾病時にお
ける涙液層の脂質および脂質 - タンパク質の相互関係に対するマイボーム腺の関与について検討することを目的にしている。
涙液油層の総説
ヒトの涙液層の主な 脂質 源はマイボーム腺である。マイボーム腺分泌物は、いろいろな種類の極性・非極性脂質のきわめて複雑な混合物であり、コレステリルエステル( CE: cholesteryl ester )、トリアシルグリセロール、遊離コレステロール、遊離脂肪酸( FFA: free fatty acid )、リン脂質、ワックスエステル( WE: wax ester )、ジエステルを含んでいる 6-9 。こうした分泌物を指す用語であるマイバム( meibum ) が用語集に記載されたのは、 1981 年以降のことである 10 。
明確に区分できる 3 つの層、すなわち糖衣(グリコカリックス)層、中間の水層、外表面の涙液油層( TFLL:tear film lipid layer )からなる現在の涙液層モデルが最初に提唱されたのは 1950 年代である 11 。糖衣層は角膜上皮を覆っており、膜結合型および分泌型ムチンを多量に含むので比較的粘性が高いと考えられている。水層は水溶性タンパク質、ムチン、塩類 に富むが、涙液油層は脂質とそれに結合もしくは挿入されるタンパク質だけでほぼ構成されている 2 。涙液油層は通常は 2 層構造を呈しており、極性脂質でできている下副層と、非極性脂質でできていて外気に触れている上層からなる 11 。この説は、 Holly 12 が最初に提唱し、後に Shine と McCulley 13 が洗練させていった。それぞれの副層には異なる役割がある。すなわち、上副層はその下に広がる涙液層の水層をすっぽり覆う厚い毛布のようなものであり、脂 質膜はその膜厚と成分に依存して水蒸気透過率が低いので、最表面の脂質成分が水分蒸発を遅らせていると考えられている 13 。非極性脂質は、水層の副相の上に広がっている時には熱力学的に不安定であり、容易に壊れて脂質小滴を形成する。そうなると涙液層の水層が剥き出しにな り、急速に蒸発することになる 13 。興味深いことに、下副層の脂質層が界面を作って、上層の安定に役立っていると考えられている。この界面において、極性脂質は疎水性の尾部を非極性脂質副 層に入れた状態で垂直方向を向いており、極性を持つ頭部を水層に触れさせていると考えられている。 Shine と McCulley 13 はこの極性脂質副層は分子 1 個から 3 個の厚みであると述べている。彼らはさらに、この極性脂質副層を構成するものはリン脂質を始めとする極性脂質であり、フォスファチジルコリン、フォスファ チジルエタノールアミン、スフィンゴミエリン( SM: sphingomyelin )、セラミド、セレブロシドなどが含まれるとしている 13 。最近になって、両親媒性脂質のもうひとつのグループすなわち超長鎖 (O- アシル )-ω- ヒドロキシ脂肪酸がマイバム成分として見つかり、これが極性脂質副層に寄与しているという説が出されている 14 。極性-非極性脂質層の厚みの実測はまだ行なわれていないが、極性脂質は全脂質分画の 5 %から 15 % 2,6 になると言われている。したがって、この極性のある界面活性物質層は分子 7 個から 20 個の厚みで、極性脂質層の上により疎水性の高い脂質があるという構成になっていると想定されている 6,15 。この推測には、(十分に可能性はあるが)異なるクラスの脂質が副層間で再配分することは考慮に入れておらず、涙液油層の全脂質成分の構成はマイバムと同 一であるという仮定に基づいている 2 。
脂質は、涙液油層に会合するクラスの分子のみではないと思われる。例えば、現在では、タンパク質が涙液油層の不可欠な要素と考えられつつあ る 2 。多くのタンパク質は表面活性化合物である。このことは、多くのタンパク質が気相液相界面に自然に密集することを意味し、水の界面張力が低下し、界面タン パク質層が形成される。バルク水相から気相液相界面へのタンパク質の移動は、概してタンパク質変性(すなわち、不可逆的構造変化―一般的にはアンフォール ディング―最終的にタンパク質の水深層への浸水を防ぐ)を伴う。これにより、タンパク質層が形成される 2 。
マイボーム腺の脂質が存在すると、タンパク質は占有する表面積について競合せざるをえない。この競合の結果、脂質層へのタンパク質の侵入 (挿入)または脂質層へのタンパク質の付着(もしくは会合)が起こる。いずれの場合も涙液層および涙液油層の表面特性に変化を来す。実際、涙液リポカリン ( Tlc )が全体の電荷や構成成分に関係なく様々な人工脂質膜と能動的に相互作用できることを Saaren-Seppaälä ら 16 が実証しており、別のグループはマイボーム腺脂質に結合したヒト Tlc が薄膜を形成することを示している。これより以前に行なわれた同様の実験でも、その他の涙液タンパク質(リゾチーム 17 やムチンなど 3 )が脂質層に侵入する可能性があることが示されている。
以上のように、涙液層の従来の 3 層モデル( Fig.1 )と 涙液油層 の 2 つの副層からなる最新の説は十分に根拠を持つ。この新しいモデルには、 涙液油層 に挿入または吸着しているタンパク質(リポカリン、 リゾチーム 、ムチンなど)と、ヒトのマイバムで最近同定された新しいクラスの脂質であり、最表面の非極性脂質副層と涙液層の水層との中間層をなす界面活性脂質副層の 形成に働いていると考えられる 8,15,18 、超長鎖 (O- アシル )-ω- ヒドロキシ脂肪酸が組み込まれていなければならない。
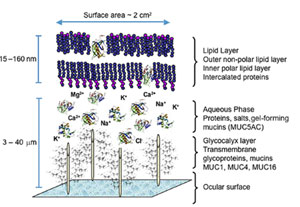
FIGURE 1. A proposed model of the precorneal tear film showing the relationship and interaction of lipid- binding proteins and the outer lipid layer.
脂質評価のための分析手法
脂質の検出と同定の技術の進歩に伴い、サンプルの質がきわめて重要となる。現代の装置では、広範囲の種類の成分を検出可能であり、その構造 について正確な情報を得ることができる。マイボーム腺の脂質の中で過去には検出や同定ができなかった多くの分析物が、たとえ微量にしか存在しなくとも、今 では見つけ出して分類することが可能である。
マイバムおよび涙液層中の脂質のサンプル採取と保存の方法
脂質サンプルの扱いと 保存は概して、 脂質化学の教科書に記載されている推奨と脂質化学企業のウェブサイト(例: http://www.avantilipids.com 、 technical support, lipid storage, and handling )で入手できるプロトコールに準じ ている 19 。サンプルがプラスチック製ないしはシリコーン製の製品に曝露または接触することを最小限にする(または防止する)ために注意を払わなければならない。ガ ラス、ステンレス鋼、貴金属類、テフロン( E.I. du Pont de Nemours and Co. , デラウェア州ウィルミントン)が、採取および保存のための素材として推奨される。脂質サンプルを保存するのに適した条件は、そのサンプルの由来を問わず、 低温 ( -80°C )、 ガス(アルゴンまたは窒素)置換、乾燥した暗所である。
マイバムと涙液層のサンプルは、次のような 3 種類の主な手技で採取可能である: ( 1 ) マイボーム腺開口部からのマイバムの弱圧・強圧による圧出 10,20-22 、 ( 2 ) 涙液水層サンプルおよびマイボーム腺開口部からのマイバムのマイクロキャピラリーによる直接採取 23 、 ( 3 ) シルマー試験紙または同様の用具による涙液水層サンプルの採取 2,24,25 。動物実験およびヒト剖検ではもうひとつの手法として、眼瞼やマイボーム腺の外科的切除も用いられるが 26-28 、 生存する ヒト有志者に対しては現実的でない。
弱圧による圧出、すなわち眼瞼の表面からの圧迫のみを行なう手法は、眼瞼の扱いが優しいので、周囲の組織によるサンプルの汚染の可能性がおそらくほとんど ない。強圧による圧出は絞り出し法とも呼ばれ、コンフォーマーなどの装置を眼瞼の後に入れて、眼瞼の前から圧を加えるものである。こ の手法により
大量のサンプル量を採取できるが、 被験者にとっては不快感が大きい。いずれの手法とも、採取されるサンプルは涙液や周囲の組織(細胞または 残屑 )で汚染される可能性がある。汚染の程度は患者および検者で 異なる可能性がある 。
サンプルが検者の皮膚分泌物 によって 汚染されることを防ぐために、検者はパウダーフリーでラテックスフリーの手袋を装用することが強く奨められる。サンプル採取の時に細隙灯を用いると、マイ ボーム腺開口部の視認性が向上し、眼瞼縁の涙液や残屑による汚染の機会を減らすことができる。
マイクロキャピラリーによる涙液層採取の手法は事実上無痛であり、下眼瞼の涙液メニスカスから涙液水層を直接採取することが可能である。涙 液分泌を刺激せずにこの手法で採取したサンプルは、一般的に 2 から 5 μL の間、すなわち数 μg の湿潤サンプルとなる 18 。マイクロキャピラリーによる手法は個別のマイボーム腺開口部からマイバムを直接に採取する際にも用いられている 22 。この手法は強圧による圧出に比べて患者の忍容性に優れ、侵襲も少ないが、採取できるサンプル量が少ない。マイバムサンプルは室温で固化する 18,29 。固化したマイバム脂質をマイクロキャピラリーから取り出すには細い金属線で押し出すか、またはクロロホルム / メタノールに単に浸すだけでもいい。マイバム採取にスパーテルを用いた場合は、サンプルを直ちに溶媒の中に保存する。マイクロキャピラリーでは、平均して 一眼あたり 15 mg のマイバムが採取される。この量はダウンストリーム質量分析による解析的分析を行なうには一般的に十分だが、タンパク質の抽出やさらなる分析を行なうため には、プールする必要がある場合もある。
患者はシルマー試験紙法を眼表面検査の 一般的項目とし て受け容れている。シルマー試験はマイクロキャピラリーによる手法ほどには患者にとって快適ではないが、その点はこの試験が サンプル採取の際に 鋭利で硬い器物を使用しないという安全性の高さで十分に補える。ただし、シルマー試験では涙液水層で試験紙を濡らさずにサンプルを採取することは事実上不 可能なため、純粋なマイバムのサンプルの採取には使えない。したがって、純粋なマイバムのサンプルの採取には圧出とマイクロキャピラリー採取またはスパー テル採取の組み合わせが適しており、マイクロキャピラリーとシルマー試験紙は涙液水層の採取に適していると思われる。脂質サンプルの採取と保存の別法と最 適法について、さらに評価を行なっていく価値がある。
薄層クロマトグラフィー
薄層クロマトグラフィー( TLC: thin-layer chromatography )は、広範囲のクラスの脂質を定量する方法として確立された手法である。しかし TLC は大量のサンプルが必要であり、そのために涙液とマイバムのサンプルの研究が困難な場合がある。検出は染色(ブロモチモールブルー)またはチャーリング( charring )で行なう 30 。それぞれの脂質の種類を同定するには、 TLC プレート の対象領域を切り取って、脂質を抽出し、化学的に誘導体化して蒸気にしてガスクロマトグラフィーにかけるか、発色団または蛍光発色団で標識して高速液体ク ロマトグラフィー( HPLC: high-performance liquid chromatography )にかけて検出する。この手法で 1976 年にウシのマイボーム腺分泌物が初めて分析された 31 。 McCulley と Shine 32 は TLC の後に UV 検出による HPLC とガスクロマトグラフ質量分析( GC-MS: gas chromatography-mass spectrometry )を用いた。脂質の定量法として TLC がいまだに頻用されるが、マトリックス支援レーザー脱離イオン化法( MALDI: matrix assisted-laser desorption ionization )や
エレクトロスプレー法といったより新しい手法が急速に受け容れられてきており、それら単独で、もしくは TLC との組み合わせで利用されている。脂質サンプルは、誘導体化または標識を行なわなくても、 MALDI とエレクトロスプレー法で分析可能である。
質量分析
質量分析は非常に感度の高い分析法であり、極微量のサンプルから検出と構造決定が可能である。数多く発表されている質量分析を用いたマイバ ム研究では、分析法として GC-MS 10,33 、液体クロマトグラフ質量分析 7,9,18,20,34 、エレクトロスプレー法 8,9,23,35,36 、大気圧化学イオン化法 7,18,20 、 MALDI 37,38 が用いられている。ウサギとヒトの涙液からはリン酸化脂質が検出されており、独特な抽出法とサンプル調整を用いた MALDI-TOF (飛行時間型)法で分析されている 38 。ヒトのマイバム脂質を MALDI-TOF を用いて検出した論文はまだない
核磁気共鳴分光法
核磁気共鳴( NMR: nuclear magnetic resonance )は、 1 H 、 13 C 、 31 P 、 17 O 、 15 N などのスピンの性質を有する原子核の量子力学的な磁気特性を測定するものである 39,40 。 NMR 分光法はサンプルを破壊しないので、より複雑な HPLC や質量分析の測定にかける前に主要な脂質クラスを定量する手法として有望である。 NMR 手法は妥当性の検証が欠かせず、バンドの割り付けは注意深く確認しなければならない。 NMR の欠点は、ミリグラムの量のサンプル が必要であることと、ワックス類の NMR 分析で選択される溶剤である重水素化シクロヘキサンにはリン脂質が溶解しないことである。したがって、 NMR 分析にはプールしたマイバムサンプルが必要となる。
赤外分光法とラマン分光法
赤外分光法の基本原理は、入射電磁波(赤外線)と同じ周波数で電子双極子の結合状態が変化すると、光が吸収されることである。吸収された電 磁波の強度と周波数から分子の振動状態の遷移が測定でき、そこから、分子レベルでの成分、環境、立体配座に関する情報が得られる。赤外分光法とラマン分光 法は補完的な手法であり、いずれも振動モードを測定するが、対称的なバンドならば一般的にラマン分光法のほうが強度 は 大きく、非対称的なバンドならば赤外線吸収スペクトルのほうが強度 は 大きい。蛍光は、ラマンスペクトルほどには赤外スペクトルに干渉しないが、ほとんどの生物組織で見られる水バンドが赤外線スペクトルの特徴を圧倒して干渉 することがある。赤外線バンドはもともとラマンバンドよりも幅が広く、解像度が低い 39,40 。
ラマン分光法と赤外分光法は、ヒトのマイバムとヒトの涙液脂質の立体配座の解明に利用されている 24,41-44 。マイバムの成分構成は涙液脂質とは異なっており、 C=C と CH 3 成分が涙液脂質より多く含まれる。マイバムと涙液脂質では立体配座も異なっている 24 。
脂質の化学特性
脂質は、構造的にも、機能的にも、きわめて複雑な分子の集合体である。化学特性による分類方法のひとつが、脂質を極性、両親媒性、非極性による分類である 45 。定義により、極性脂質は比較的水溶性である。このグループには、短鎖脂肪酸、ヒドロキシル化された脂肪酸、ヒドロキシセラミド( OH-Cer: hydroxy-ceramides )、モノアシルグリセロール( MAG: monoacyl glycerol )、グリコシル化された脂質、リン脂質などが含まれる
45 。これらの脂質は親水親油バランス( HLB: hydrophilic-to-
lipophilic balance )が比較的高い。 HLB は、極性(水性)と非極性(油性)の副相間で溶解している分子の分配率を表わす客観的な物理化学パラメータである。したがって、油中水型エマルジョンにお いては、極性脂質は水層の副相内で凝集する。それに対して非極性脂質は水に溶解しない。このグループの典型的なメンバーとしては、炭化水素、超長鎖アシル セラミド、 WE 、 CE 、トリアシルグリセロールがある 45 。これらの脂質の親水親油バランスはきわめて低く、油中水型エマルジョンの油相のほうに容易に分配する。炭素鎖の長さ、構造における親水基の数と種類(水 酸基、アミノ基、カルボキシ基)、不飽和の程度、シス・トランス・イソ型異性体など、脂質の溶解度に影響する要因が多数存在することから、これら 3 つのグループの境界線は曖昧である 45 。
複合脂質の形成は、主としてエステル化とアミド化といった縮合反応を通じて生じる。エステル化反応は主要な複合脂質のほぼすべて( WE 、 CE 、アシルグリセロール、リポ多糖)の生合成に関わっており、アミド化が形成に関与する脂質は脂肪酸アミド( FA: fatty acid amide )、 Cer 、 SM 、セレブロシドなどそれより少ない 45 。これらの反応は可逆的である。すなわち複合脂質は in situ で酵素的または非酵素的に加水分解され、その過程で、複合脂質はより単純な脂質と、グリセロール(アシルグリセロールの場合)や炭化水素(リポ多糖の場 合)といった脂質以外の物質の混合物に戻る 45 。
涙液層と 涙液油層 に関係する脂質の変質の種類として 2 番目に重要なものが、脂質の酸化である。脂質酸化の必須条件のひとつが脂質構造に二重結合が 1 つ以上存在することである。二重結合は酵素的または非酵素的に酸化することができる。多くの脂質には酵素的または自然発生的な変化で誘導される異性体があ る。高温、紫外線、酸化が異性体形成の一般的な原因である。これらの反応は、脂質とその混合物の物理化学性状を必然的に変化させる。というのも、 ( 1 ) 加水分解産物が一般的に元の複合脂質よりも水溶性が高く、 ( 2 ) 脂質の(過)酸化産物も酸素付加や短くて親水性の高い切断産物の形成により親水性が高くなり、 ( 3 ) 異性体形成によって脂質パッキングと脂質の物理性状(融点、沸点、密度など)が影響を受けるためである 45 。
現在では、ウェブサイトで脂質の情報を容易に入手することができる。 lipidmaps.org 、 lipidli-brary.co.uk 、 cyberlipid.com 、 hplc-ms.byrdwell.com 、 lipid- banks.jp などのサイトが推奨される。最近ではリピドミクスの概要も 発表 されており、これら生体分子の分子的構成が明確に説明されるようになっている 46 。
涙液層の脂質
正常のマイバム
マイバムの生化学的性質の解明が始まったのは 1897 年であり 47 、 Orlando Pes がその脂質の性状を確認し、脂肪、 FFA 、コレステロールに富むという説を述べた。その数十年後に Linton ら 30 、 Andrews 48 、 Ehler 49 がそれぞれ、マイバムには中性脂肪、ステリル、 WE が豊富にあることを示した。続く研究によって、アシルグリセロール、セラミド、リン脂質などの極性脂質がマイバムに存在することと、マイバム成分のほとん どがいろいろなクラスに属する非極性脂質であることが報告された 28,31,50-52 。極性脂質は少数だが、涙液層の安定と崩壊においてきわめて重要な役割を持っている。この トピック に関する総論が複数公表されており 53-56 、国際ドライアイワークショップも最新の総説
を出している 57 。今日までにさまざまな手法で明らかにされた脂質のすべてを Table.1 にまとめた 7-10,13-15,20,23,29,32-34,36,47-50,52,56,58-74 。
マイバム脂質の中でもっとも頻繁に検出されるクラスはおそらく遍在する WE と CE である。この 2 種類を合わせて、マイバム脂質の 60 %程度を占めると考えられている。 WE と CE はマイバムの中でもっとも疎水性の高い成分であり、その親油性に匹敵するのは炭化水素のみである。
近年、ヒトの正常マイバムの WE の特性が HPLC-MS n で明らかにされている 7,14,18,20 。多数の WE の 種類が検出されており、もっとも多く存在するものは 18 から 30 個 の炭素を持つ超長鎖飽和脂肪アルコールの C18:1- 脂肪酸由来のエステル体であった 7,18,20 。その後の研究により、記載される WE の種類がさらに増え、 C18:2 、 C18:3 、 C18:4 脂肪酸ファミリーなどが含まれるようになった。これらの脂肪酸はエステル化して、 C18:1 脂肪酸同様に、同じ長さの超長鎖脂肪アルコールになる。多価不飽和 WE は一価不飽和 WE に比べて少ないが、それでもマイバム成分として顕著に存在している。これらの多価不飽和 WE を詳しく分析すると、これらのファミリーの各メンバーの多くが、複数の異性体の形で存在していることが明らかになった。異性体は二重結合部位のシス・トラ ンス幾何学異性体である 可能性が高い 17 。例えば、 m/z 641 のある物質( C26:0 アルコールと C18:4 ステアリドン酸のエステル、 FAl:FA 、 C26:0 : C18:4 )は 4 種類の異性体の形で存在しており、 C26:0 : C18:3 は 3 種類、 C26:0 : C18:2 は 2 種類の異性体で存在しているが、 C26:0 : C18:1 は 1 種類の異性体のみで存在している。こうした観察データから、ヒトのマイバムに含まれる個々の WE 種の数は全部で 100 を超えることが想定される。直接注入型 ESI による WE の分析では、主に C18:1 が見つかるが、 C16:1 もかなりの量が存在する 8 。もうひとつのグループ( CE )に属するマイボーム腺脂質も同様に複雑である 8,15 。大部分の脂肪酸が中間的な長さの C18:1 から C18:4 に属する WE とは異なり、マイバムで検出される CE には超長鎖の飽和または不飽和脂肪酸もあり、その長さは炭素 18 から 34 個までの幅がある。ヒトのマイバムでは CE が 20 種類以上見つかっている。主体をなす種類は炭素 22 から 30 個 の脂肪酸による CE である。マイボーム腺の全 CE の中でのオレイン酸由来の C18:1CE (その他の組織や生物で一般的な CE のひとつ)のモル比は 5 %に満たない。遊離コレステロールもマイボーム腺のステロールとステリルエステルにおいて 0.5 %未満しかないようである 15 。
ヒトのマイバムにおけるリン脂質の相対量については、異論が多く未解決であるが、リン脂質の量は以前考えられていたほどには多くないようである 6,7,13,20,22,75,76 。リン脂質の定量と同定における意見の不一致の原因はおそらく ( 1 ) サンプル採取手法のばらつき、 ( 2 ) マイバムサンプルが涙液水層で汚染される程度のばらつき、 ( 3 ) 器具とそれに関連する手法の差にあると考えられる。しかしマイボーム腺の分泌メカニズムが全分泌型であり、細胞膜がリン脂質に富むことを考えれば、リン脂 質は少なくとも最初は腺からマイバムに分泌されると思われる。こうした極性脂質の検出と、性質および想定される変異の程度の評価を行なう能力は、データ分 析法に依存して実験グループ間で異なっている可能性がある 9,77-79 。脂肪酸のアシル 置換 基や鎖長の不飽和など、それぞれのリン脂質分子種の感度に影響を与える可能性がある要因がいくつかあるが、中でももっとも顕著な要因がイオン化効率であ り、これはそれぞれのリン脂質クラスの極性頭部基に依存する。主要なクラスがイオン源に存在している場合には、イオン抑制効果によって少量しかない成分の 検出が難しくなる。結局、リン脂質の中には陰イオンの状態のほうが容易に検出できるものも |