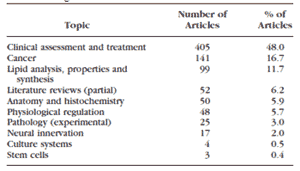
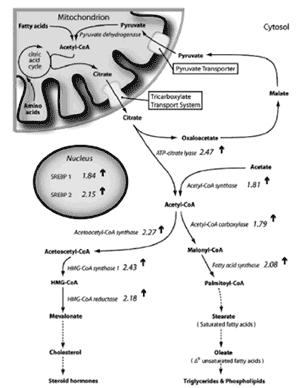
• 脂肪酸生成酵素、 HMG-CoA 生成酵素、 HMG-CoA 還元酵素、メバロン酸キナーゼ、ホスホメバロン酸キナーゼ、メバロン酸ピロリン酸デカルボキシラーゼ、イソペンテニルピロリン酸イソメラーゼ、ゲラニルゲ ラニルピロリン酸生成酵素、スクアレンエポキシダーゼ、ラノステロールデメチラーゼ、 Δ 7 - ステロール還元酵素はすべて、コレステロール、脂肪酸、性ステロイド、脂質の合成を開始進行させるのに中心となる酵素である 232,233 。その他の組織でも脂肪酸合成酵素はアンドロゲンで制御されることが分かっており 214,218 、マイボーム腺上皮組織で発現する( Richards SM, et al. IOVS 2002;43:ARVO E-Abstract 3150 )。
• 脂肪酸輸送タンパク質 4 は長鎖および超長鎖脂肪酸の細胞内取り込みと代謝を促進する 234 。
• 超長鎖脂肪酸伸長様酵素タイプ 1 とタイプ 3 は超長鎖脂肪酸とスフィンゴ脂質の組織特異的な合成を増大させる 233,235 。これらのタンパク質は、ウサギのマイボーム腺において総脂質分画の中でのアンドロゲン誘導性の長鎖脂肪酸の増加に関与している 223 。
• モノグリセリドリパーゼは、トリグリセリドとモノグリセリドの脂肪酸とグリセロールへの加水分解を促進する 236 。
• ATP 結合カセットファミリーに属する Abca1 と Abcd3 は、膜外および膜内で様々な分子を輸送する。 Abca1 は脂質除去経路においてコレステロール流出ポンプとして働き 237 、コレステロール配分の中心的制御因子として機能している 238,239 。 Abcd3 は脂肪酸や脂肪アシル -CoA のペルオキシソーム内への輸送を制御する 240 。
• オキシステロール結合タンパク質様 1A 、ステロール担体タンパク質 2 、リポカリン 3 、フォスファチジルコリン転移タンパク質はリン脂質の結合や輸送に関与している 233 。
• SREBP1 は脂肪酸合成経路を促進し、 SREBP2 はコレステロール合成経路を増強する 241-244 。
• 多数の遺伝子がペルオキシソーム増殖剤活性化受容体のシグナル経路に関与しており、これは、脂質生成、ケトン体生成、脂質輸送、脂肪酸の輸送と酸化を調節している 98 。
アンドロゲンはさらに、マイボーム腺の内分泌制御においてきわめて重要な遺伝子群を調節する 81 。例えば、テストステロンは次のものの mRNA レベルを上昇させる:
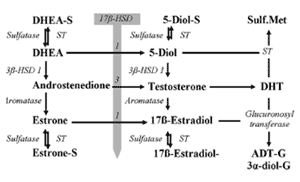
• 17β-HSD7 : 17β- ケトステロイドとそれに対応する 17β- ヒドロキシステロイドの相互変換を制御する酵素 245 。この酵素の活性は、末梢組織のすべての活性型アンドロゲン・エストロゲンの代謝においてきわめて重要であり 245 、副腎前駆物質からのマイボーム腺局所でのアンドロゲン細胞内分泌合成に関わっていると思われる。
• インスリン様増殖因子 1 :多面効果を持つタンパク質で、皮脂腺細胞の DNA 合成と分化を刺激する 147 。
• エストロゲン受容体( ER: estrogen receptor ) β :前立腺においてアンドロゲンで発現が亢進する受容体で 246 、 ERα の活性を抑制していると思われる 247 。
• 11β-HSD1 :コルチゾールから不活性代謝産物であるコルチゾンへの変換を触媒する酵素 233 。
興味深いことに、テストステロンは角化に関与する遺伝子 81,135 や、レチノイン酸生合成を促進する酵素であるレチナール脱水素酵素 3 の遺伝子 81 の発現を低下させる 233 。またアンドロゲンは、エンドソーム、ゴルジ装置、小胞体、ライソソーム、プロテオソーム、核、ミトコンドリアといった細胞内のさまざまな場所で、タンパ ク質の選別、輸送、加水分解に関わる遺伝子の発現を刺激する 81 。
以上の知見に加え、雌マウスのマイボーム腺ではアンドロゲン処置によって 1000 個を超える遺伝子の発現が影響を受けることが分かっている 135 。雌の組織のそうした遺伝子の多くが、テストステロンによって調節されており、雄のマイボーム腺においてアンドロゲンで制御されている遺伝子と同じもので ある。しかし中には、性特異的な様式で制御されている遺伝子もある 135 。
アンドロゲンはマイボーム腺の構成成分である脂質に、そしておそらくはタンパク質にも影響を及ぼす。睾丸 摘出によってウサギのマイボーム腺の脂質の分析結果が顕著に変化するが、 19- ノルテストステロンの局所または全身投与を 2 週間行なうと、(プラセボに比べて)脂質プロファイルは無処置動物のプロファイルへ回復し始める 223 。さらに、ヒトのマイボーム腺上皮細胞においてアンドロゲンのシグナル機構が secretogloblin の発現も制御しているとも考えられている 248 。このタンパク質は涙液層に分泌されて、リポカリン様機能を果たしているのかもしれない 248 。
マイボーム腺に対するアンドロゲンの作用のすべてが古典的な受容体を介して行われているのかどうかについ ては不明である。その作用には腺の膜受容体への結合、シグナル伝達カスケードの刺激、それによる遺伝子転写の変化も関与している可能性がある 249,250 。さらにアンドロゲンは、マイボーム腺に影響を与える視床下部ホルモンおよび下垂体前葉ホルモンの分泌を制御することで、間接的に作用する可能性もある。
アンドロゲン欠乏の影響と治療 アンドロゲンがマイボーム腺機能に 影響を持つことから、閉経時(卵巣と副腎からのアンドロゲン分泌の減少) 251 、男女の加齢(アンドロゲン総量の減少) 196,251 、自己免疫疾患(シェーグレン症候群、全身性エリテマトーデス、関節リウマチなど) 201,252-253 、完全型アンドロゲン不応症候群( CAIS: complete androgen insensitivity syndrome 、アンドロゲン受容体が機能しない女性) 254,255 、抗アンドロゲン薬(前輪腺の肥大や癌に対して) 256 などで見られるアンドロゲン欠乏によって MGD 、マイボーム腺分泌物の脂質プロファイルの変化、涙液層安定性の低下、および蒸発亢進型ドライアイが起きるのではないかという説が唱えられている 257-260 。
抗アンドロゲン治療を受けている患者のマイボーム腺は、対照群に比べると、開口部異形性(導管上皮の異常 増殖と角化として定義される病態) 261 、分泌物の質の低下、分泌物の中性脂質プロファイルの顕著な変化、重症例に相当する形態変化がある 258,260 。こうした脂質の変化の大部分は両眼に等しく現われるか全く現われないかであり、脂肪酸パターンに特徴的な変化が現われる 260 。また、患者では眼表面の自覚症状(光過敏、眼の疼痛、霧視など)が現れるのが多いのと同様に、涙液層(残屑、異常メニスカス、不安定など)、結膜(瞼結 膜充血、下部染色など)、角膜(染色など)、眼瞼(瞼縁後部の不整、たるみ、 collarette など)の異常の頻度が非常に高い 258 。こうした知見に沿うもうひとつの知見が、テストステロンレベルを下げるための酢酸ロイプロリドの投与で一部の患者にみられる眼の副作用や霧視と関連があ るというものである 262 。これらの結果は、前立腺肥大の治療薬とドライアイ疾患との間に有意な関連があることの説明になる 263 。
CAIS 患者におけるアンドロゲン受容体機能不全にはドライアイの自覚症状と他覚所見が有意に多いということも分かっている。この特殊な患者群は年齢をマッチング させた男女の健常者に比べると、マイボーム腺開口部の異形性と瞼縁後部の不整が有意に多く見られ、マイボーム腺分泌物の質も低下している。さらに CAIS 患者は、健常な男女からなる対照群に比べて、マイボーム腺分泌物の中性および極性脂質のパターンに顕著な変化が見られる 257,259 。正常な加齢過程でもマイボーム腺分泌物の質の有意な低下とマイボーム腺開口部の異形性の有意な増加が見られる 105,264 。マイボーム腺分泌物の極性脂質と中性脂質のいずれもが、加齢により有意に変化する 105,264 。この所見は、 37 歳と 70 歳の患者を比較して得られたものであり、男女とも 30 歳代から 70 歳代の間でアンドロゲンが劇的に減少する 251 。
こうした知見の他にも、自己免疫疾患ではないドライアイと MGD の患者はアンドロゲンが不足していることや 265 、アンドロゲン前駆物質である DHEA をヒト、ウサギ、イヌに局所投与すると、マイボーム腺脂質の産生と分泌が刺激され、涙液層破壊時間( TBUT: tear film break-up time )が延長することも分かっている 266 。さらに、テストステロンの血清レベルの低下が、ドライアイの女性患者で多く見られ、眼症状の主観的重症度と相関していることと 267 、閉経前・閉経後の女性においてそれぞれ血清テストステロン濃度がマイボーム腺分泌物の量およびマイボーム腺開口部の径と相関していることが研究で示され ている( Suzuki T, et al. IOVS 2007;48:ARVO E-Abstract 434; Suzuki T, et al. IOVS 2008;49:ARVO E-Abstract 92 )。しかし、血清テストステロンレベルはヒトの総アンドロゲン量の中でほんのわずかな割合(女性で 0.2 %未満)を占めているに過ぎず 268 、卵巣活動の指標としての意義以外の価値はまったくあるいはほとんどないため 251,268,269 、後者の研究の結果の意義は不明である。女性ではアンドロゲンの大部分もしくは
全部(閉経前は 75% 、閉経後は 100 %)が、副腎の性ステロイド前駆物質( DHEA と DHEA サルフェート)を基に末梢組織で生成される 269 。ヒトの総アンドロゲン量の妥当で信頼性のある推計値はおそらく、抱合ジヒドロテストステロン代謝産物(グルクロン酸アンドロステロンとアンドロスタン -3α,17β- ジオール - グルクロン酸)の血清濃度のみだろう 251,270,271 。これらは身体全体の細胞内分泌産生量と末梢組織のアンドロゲン代謝産物との総量を反映している 269 。
以上の知見をまとめると、マイボーム腺はアンドロゲンの標的器官であり、アンドロゲンはこの組織の脂質産 生を促進し、角化を抑制する。また、アンドロゲン欠乏により MGD と蒸発亢進型ドライアイが引き起こされる可能性がある。こうしたアンドロゲン欠乏と MGD および蒸発亢進型ドライアイとの明らかな相互関係は、男女のドライアイの自覚症状と他覚所見がアンドロゲンの局所・全身投与で軽減するという報告の説明に なり得る 266,272-278 。アンドロゲン欠乏患者のこうした内分泌のバランス異常を軽減する取り組み(アンドロゲンの点眼など)が、 MGD とそれに伴う蒸発亢進型ドライアイの治療に有益であることを証明するかもしれない。その可能性に一致する臨床試験の結果が得られており、 MGD をテストステロン点眼で治療すると、マイボーム腺分泌物の質が向上し、眼の不快感が減少するであろうことが示された( Schiffman RM, et al. IOVS 2006;47:ARVO E-Abstract 5608 )。
エストロゲン
エストロゲンによる皮脂腺の制御 さまざまな動物種においてエスト ロゲンは皮脂腺の大きさ、活性、脂質産生量を有意に減少させる 145,181,192,279-283 。実際には、エストロゲンはかつて皮脂産生を抑制するプロトタイプ薬と呼ばれており 282 、皮脂腺機能を弱め皮脂の分泌を抑えるための治療法としてエストロゲン治療が数年間用いられていたことがあった 180,181,280,281,284,285 。
このホルモン作用について提唱されているメカニズムは、エストロゲンが皮脂腺細胞内のリソソーム酵素の放 出を誘導して早期の細胞破壊を起こさせ、皮脂の分泌量を減らすというものである 282,286 。その他に言われているメカニズムは、エストロゲンがテストステロン取り込みを低下させて、テストステロンからジヒドロテストステロンへの変換を阻害し、 皮脂腺のアンドロゲン作用に拮抗するというものである 279,282,283 。事実、エストロゲンは皮脂腺へのアンドロゲン作用を減少させる治療の主力として挙げられている 180 。エストロゲンのこうした抗アンドロゲン作用は用量依存性であり、生理的レベルのアンドロゲンによる治療で打ち消される 181,280 。
興味深いのはアンドロゲン治療によって皮脂腺のエストラジオール結合部位の数が有意に減少することであり 177,286 、この 2 つのホルモンは互いに受容体を変化させる 283 ことで皮脂腺において拮抗している。さらに、アンドロゲン作用の中には、低レベルのエストロゲンに依存しているものもあると考えられている 287 。
エストロゲンによるマイボーム腺の制御 マイボーム腺にはエ ストロゲン受容体の mRNA とタンパク質が存在し 230,288,289 、卵巣摘出したマウスにエストロゲンを投与すると、腺の形態に特徴的な変化が現われる 290 。エストラジオール -17β もマウスのマイボーム腺において 200 個近くの遺伝子の発現を制御している 291 。例えば一部を挙げると、チロシンキナーゼ関連(線維芽細胞増殖因子受容体 1 )、免疫因子(イ
インターロイキン 1 受容体 II 型)、細胞外マトリックス成分(マトリックスメタロペプチダーゼ 3 )、ステロイド生成(エストロンを生理活性のあるエストラジオールに変換する 17β- ヒドロキシステロイド脱水素酵素 7 292 )、プロラクチン活性(プロラクチン受容体)、脂質代謝などに関係する遺伝子がある。
エストロゲンは脂質動態に関係する特定の遺伝子(形質膜にフォスファチジルコリン 293 を補給するフォスファチジルコリン転移タンパク質 291 など)の発現を増加させ、リパーゼの 1 種であるカルボキシルエステラーゼ 3 294 などの他の遺伝子の発現を抑える 291 。このようなホルモン作用は 17β- エストラジオールがマイボーム腺の脂質産生を促進することを示唆しているが、エストロゲンの作用のほとんどがこの結論に矛盾することが分かっている。 17β- エストラジオールはむしろ脂質産生において全体的にマイナスに影響するようである。例えば、エストロゲンは脂質や脂肪酸の異化に関与する複数の遺伝子の発 現を刺激し(抗脂質産生遺伝子の STAT5A 295 )、脂質の生合成、動員、処理、膜輸送に関与する遺伝子を抑制する 291 。
後者のこうした抗アンドロゲン作用とともに、エストロゲンが皮脂腺に及ぼす全体としての影響を考えると、エストロゲン治療はマイボーム腺の脂質合成を低下させ、 MGD および蒸発亢進型ドライアイを促進すると考えるのが合理的である。この考えの根拠を以下に挙げる:
• 25,665 人の閉経後女性を対象にした疫学調査で、エストロゲン補充療法を受けている集団はその治療を受けたことがない集団に比べて、重度ドライアイ症状を保有する率が高く、ドライアイ症候群の臨床診断される率が高かった 296 。
• 44,257 人のドライアイ女性患者を対象にした調査で、同時に見られる条件としてもっとも多かったもののひとつが、エストロゲン補充療法の使用であった 297 。
• 2 つの研究におけるエストロゲン治療によって、涙液層不安定、異物感、コンタクトレンズ( CL )不耐、眼表面の乾燥が引き起こされた 298,299 。
マイボーム腺の他の遺伝子は 17β- エストロゲンで抑制されるが、テストステロンで刺激される 291 。そうした遺伝子にコードされているものとしては、酸性でシステインに富む分泌糖タンパク質(細胞増殖を制御する)、血管内皮増殖因子 A (細胞遊走を促進する)、カテプシン K (細胞外マトリックスを分解する)、マトリックスメタロペプチダーゼ 3 (フィブロネクチン、ラミニン、ゼラチン、コラーゲンを分解する)などがある。これらの遺伝子が、マイボーム腺の細胞の成熟、遊走、全分泌型分泌に関与し ている可能性がある。もしそうならば、これらのホルモン反応はエストロゲンの抗皮脂腺作用とアンドロゲンの皮脂腺促進作用と矛盾しないことになる。
プロゲスチン
プロゲスチンによる皮脂腺の制御 かつては、プロゲスチン治 療により皮脂産生が有意に増加するので 146,181,300 、男性のアンドロゲンのアナログ物質であるプロゲステロンが女性の皮脂腺分泌を制御する刺激ホルモンであると考えられていた 181 。しかし初期の研究とは食い違ってその後の研究では、プロゲスチン投与は皮脂腺の分泌量に影響がないことが分かってきており 191,300 、このホルモンによって局所のアンドロゲン代謝と活性が抑制されることで、むしろ皮脂腺機能が低下することを報告する研究もある 301-304 。こうした知見の食い違いに対する説明のひとつが、各種の皮脂腺への
プロゲスチンの作用は用量、内分泌環境、被験者の性別に顕著に影響されるからというものである 302,305-307 。
プロゲスチンによるマイボーム腺の制御 マイボーム腺にはプロゲス テロン受容体の mRNA とタンパク質が存在し 230,288 、プロゲステロン曝露に対して明らかな形態変化を伴って反応する 290 。エストロゲン補充療法にプロゲスチンを追加すると、エストロゲンに起因するドライアイ症状が顕著に低減する 296 。これはマイボーム腺活動性への陽性の影響の表われと見なせる。プロゲスチンの腺への作用と眼表面症状との間の明白な関連性については今後明らかにする必 要がある。
最近の研究によれば、プロゲステロンはマウスのマイボーム腺の遺伝子発現に顕著に影響する 291 。そのほとんどの遺伝子がプロゲステロンで発現が抑制されるものであり、例としては、免疫過程、糖新生、エネルギー伝達に関連する遺伝子がある 291 。もっとも顕著な効果は、リボソームの生合成、会合、構造に関連する遺伝子のすべてが抑制されることで 291 、このことから、プロゲステロンはマイボーム腺のタンパク質、巨大分子、細胞内の生合成に対して全体として抑制的影響があると考えられる。
エストラジオール -17 βとプロゲステロンを組み合わせて投与すると、マウスのマイボーム腺において 300 個を超える遺伝子の発現に対して有意に影響した 291 。ほとんどの分子生物学的反応は単独エストラジオール治療と単独プロゲステロン治療とでそっくりであった。しかし、細胞内局所オントロジーに関与する遺伝 子の独自の発現亢進など、そうでないものもあった 291 。このプロゲスチン/エストロゲン複合反応の理由は不明である。
マイボーム腺の性差における性ステロイドの関与
まとめると、性ステロイドはマイボーム腺の機能に対して顕著な影響力を持っており、個々のステロイドに よって MGD と蒸発亢進型ドライアイを予防または促進する場合がある。さらに、性ステロイドのそれぞれ異なる作用が、他の皮脂腺 172,175,181,189,308 に対するのと同じくマイボーム腺の形態、遺伝子発現、中性・極性脂質プロファイル、分泌量に見られる既知の性差にも関与している可能性がある 44,76,105,120,259,263,264,290,309-312 。この提唱に一致する 知見 として、マイボーム腺の遺伝子発現における性関連の変動のほぼ 30 %がアンドロゲンによるものという観察がある 76 。さらに、性特異的なアロマターゼ活性もマイボーム腺の性差で役割を持っている可能性もある( Liu S, et al. IOVS 2007;48:ARVO E-Abstract 5657 )。
オールトランスレチノイン酸のマイボーム腺への作用
オールトランスレチノイン酸は皮脂腺の増殖、発達、脂質産生を低下させ、眼瞼結膜炎の原因になる 313-318 。また、皮脂腺細胞の中で 13-cis レチノイン酸(イソトレチノイン)に変換され 319 、マイボーム腺導管の肥厚と角化 320,321 、マイボーム腺腺房細胞の変性と壊死 322 、腺房周囲の線維化、マイボーム腺組織の脂質量の 減少 320,321 を引き起こすことが動物モデルで知られている。イソトレチノインは重症の痊瘡の皮膚科治療薬として数十年にわたり革命を起こした薬物である 323 。
ヒトでは、 13-cis レチノイン酸の投与によって眼瞼結膜炎、異常なマイボーム腺分泌、マイボーム腺萎縮、 TBUT の短縮、涙液層の浸透圧亢進、ドライアイの徴候と症状が生じる 324-331 。実際、このレチノイン酸派生物は MGD と蒸発亢進型ドライアイを増悪させる 330,332,333 。
レチノイン酸のこの作用に関 わる主なメカニズムとしては、アンドロゲン受容体の mRNA およびタンパク質の抑制と 334,335 、レチノール脱水素酵素 -4 の抑制によって局所のジヒドロテストステロン産生を減少させること 336 などがある。レチノイン酸のマイボーム腺への影響の解明は非常に重要である。というのも、この物質は眼の周辺に用いるアンチエイジング化粧品の多くの中心 成分であり、ヒトは年齢を重ねるにつれ、ドライアイを発症しやすくなるからである 263,312 。
増殖因子などの物質および疾病のマイボーム腺への影響
他 の さまざまな物質と疾病が、マイボーム腺の生理と病態生理に影響を与えることが分かっている。その例としては次のものがある:
• 上皮成長因子( EGF : epidermal growth factor )とウシ下垂体抽出物。これらはヒトの不死化したマイボーム腺上皮細胞の増殖、そしておそらくは分化も促進する( Liu S, et al. IOVS 2009;50:ARVO E-Abstract 3669 )。 EGF は、ウサギのマイボーム腺 337 とヒトの皮脂腺 155 の上皮細胞の分化を in vitro で刺激することも知られている。癌患者への EGF 阻害剤の治療に伴って MGD が現われる( Joshi J, et al. IOVS 2008;49: ARVO E-Abstract 2363 )。
• エピネフリンの点眼投与。これは導管上皮の過角化を誘導し、マイボーム腺の角栓と拡張を引き起こす 338 。
• ω-3 脂肪酸。この摂取量はヒトのマイボーム腺分泌物の極性脂質プロファイル 120 と飽和脂肪酸成分 119 の変動に相関している。また、 ω-3 脂肪酸の摂取量の減少が、シェーグレン症候群の女性患者で見られ 339 、この種の患者は典型的に MGD を併発する 340 ( Krenzer KL, et al. IOVS 1999;31:ARVO Abstract 2864 )。
• 視床下部性性腺機能低下症。これは 35 歳男性の閉塞性 MGD 患者で見つけられた 162 。眼瞼の状態はアンドロゲン欠乏によるものだと医師は述べている 162 。
• アルドステロン。おそらく、ヒトの皮脂腺には鉱質コルチコイド受容体が存在することによる 341 。しかしヒトのマイボーム腺組織でアルドステロンの標的になり得るものは知られていない。アルドステロンには、ウサギのマイボーム腺の中性脂質およびガン グリオシド組成に対する作用はない 342 。
• 多発性内分泌機能低下症(アジソン病や副甲状腺機能低下症)。これには重度の MGD が伴う 343 。この知見を考えると、マイボーム腺に副甲状腺ホルモン受容体の mRNA が存在する( Table 3 )ことは特に興味深いが、この受容体の mRNA が翻訳されるのか、機能するのかについてはまだ分かっていない。
また、ヒトのマイボーム腺は他の皮脂腺に同じく 200,344-347 、脂質だけでなく、タンパク質も分泌する 134 ことも興味深い。こうしたタンパク質が涙液層の安定性や眼表面の状態に対して有意に影響を及ぼしている可能性がある 134 。しかしこのタンパク質分泌過程の制御や意義については解明が待たれる。
健常・疾病の両方の場合でマイボーム腺を制御する生理的メカニズムの理解を深めるには、(いろいろある中でも)内分泌、神経、栄養、増殖因子がこの組織に及ぼす影響の性質と程度を明確に示す研究が必要であることは間違いない。
病態生理と病理
過角化
過角化は、閉塞性 MGD の主な原因であり、変性した腺の拡張と炎症のない萎縮を起こさせる。これがマイボーム腺の典型的
な病理ということは、この腺の発生段階を見れば意外ではない。過角化は、導管上皮 で生じる完全角質化(角化)の発生上阻害していたものが取り除かれたことを表わしている可能性がある。なぜなら、角化は新規 に 獲得した因子ではなく、様々な内因性・外因性の要因により生じるからである。上皮の角化やマイボーム腺の閉塞を増加させる因子には年齢 50,87,348 、ホルモンバランス異常 257,291,349 、薬物と化学物質の毒性作用 350,351 、マイボーム腺脂質の分解産物 41,352,353 、エピネフリン点眼薬といった外部要因の影響 354 から CL 装用 355 まで幅広いものがありうる。
Korb と Henriquez 3,356 が初めて記述した過角化による閉塞性 MGD の患者は、眼の乾燥感を示唆する軽微で一過性の症状しか呈していなかったが、 CL 不耐のために症状が発現するようになった。患者のマイボーム腺 の拇指による圧出 で、開口部が閉塞していることを確認し、角化物の塊が落屑上皮細胞と濃縮したマイバムで構成されていることを明らかにした。圧出と角栓除去後は、涙液層が 正常化し、 CL 不耐が消失した。閉塞した腺の組織像では、細胞残屑と皮脂物質で中央導管が拡張していることが確認された 3,356 。
その後、症候性ドライアイで導管開口部が乾固しており、きわめて粘稠なマイバムが圧出される患者から採取 したマイボーム腺の組織学的検査によって、角化増大による排出導管の閉塞所見が確認された。腺の内部は、角化増大により導管の閉塞と拡張および嚢胞状変性 と、マイボーム腺分泌細胞が消失して腺房上皮の扁平上皮化生を生じていた( Fig.18, 19 ) 357 。こうした変化は、炎症性白血球が存在せずに起こっていた。このことから、観察された腺房の変性と萎縮は、導管系の拡張後に生じ、腺の閉塞の結果マイボー ム腺細胞が消失したために二次的に分泌低下を続発したと考えることができる。同様の病理は、外科手技または新生物による閉塞で嚢胞状拡張をした腺でも観察 される。これらの症例でも、導管系の拡張が腺房の萎縮とともに報告されている 31 。この病態での導管系の拡張は、閉塞性 MGD の所見に比べると、かなり目立つようである 357 。 72 例の検視解剖のマイボーム腺を対象にした大規模な組織病理学的研究により、導管と腺房の嚢胞状拡張が確認され、これら病理学的変化がある割合は症例の 34.7 %( 25 例)と報告された 358 。現在では、閉塞性 MGD ( Fig.18,19 )がこの疾患のもっとも多く見られる形態のようである 3,4,10,357-362 。
MGD の動物モデル
自然発生もしくは導入による MGD 動物モデルが何種類か同定ないし開発されている。こうしたモデルは 、同様に、 この病態の基礎にある病態生理メカニズムの探索に非常に有益である。ウサギ 320,338,354,363,364 とサル 365 において、エピネフリンの局所投与 338,354,363,364 、イソトレチノインの全身投与 320 、ポリ塩化ビフェニル中毒 365 によって特徴的な過角化が誘発されるモデルが作製されている。 Fig.20 にあるように、これらウサギとサルのモデルの一般的な組織病理 検査 所見で、内腔に角化物が詰まった導管の異常な拡大が見られる。また、導管開口部の上皮は典型的 に 角化が高度であり、閉塞している( Fig.20B )。現時点では、開口部自体の過角化が、導管上皮と同じく、閉塞性 MGD で見られる内腔の角栓形成に強く寄与していると考えられている。
マウスの MGD および腺過形成( MGH: meibomian gland hyperplasia )のモデルも開発されている。これらは自然発症、もしくは 免疫付与、変異、遺伝子導入、ノックアウト技術に |